涡虫一类的扁虫能够将自己的身体分裂为片段,并由这些片段再生出新的完好的涡虫。科学家最近揭示了调控这种裂殖发生频率的分子机制。
要了解安排和器官怎么再生,首要就要弄清楚特定的机制和因子怎么在空间和时间上调控细胞和安排。涡虫被切开下来的片段能够重新长出缺失的身体并形成完整的虫体,因而被广泛应用于再生研究。涡虫同时具有自我切开的功能,被称为裂殖,它们延展身体并缩小尾部安排,使身体后部开裂并成长出克隆。裂殖发作与否取决于亲虫的巨细,但其间的分子和细胞进程尚不明朗。Arnold等人在《自然》发表文章,报道建立了一种可靠的能够诱导涡虫Schmidtea mediterranea裂殖的办法,并且表明触及Wnt和转化生长因子β (TGF-β)蛋白的细胞信号转导通路是调节这一进程的要害。
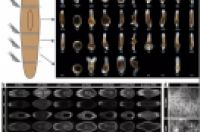
再生片段和裂殖亲本的形态和/或内部组织的分析。 图片来源:Arnold et al。(Reference 1)
Wnt信号通路在发育和细胞分化中起着决定性作用,并与许多疾病密切相关[2]。Wnt蛋白具有高度的多样性,只存在于动物中,由细胞分泌并通常附着于一条脂链上。它们通过与不同家族的受体蛋白结合,激发不同的下游细胞信号级联来调控细胞质因子的水平,这些细胞质因子可以控制基因表达,进而控制细胞功能。虽然近年来我们对于Wnt信号通路如何影响组织形成的理解有了很大的进步,但对于这一过程与特定组织功能之间的关系仍不甚了解。
过去关于涡虫的研究已经描绘出了组织形成的分子构架,自发Wnt蛋白浓度梯度控制沿动物长度方向的组织形成(即前后轴方向),TGF-β浓度梯度控制由上到下方向的组织形成(即背腹轴方向)。有研究指出涡虫裂殖受沿前后轴方向的代谢活动梯度、定位分子或神经激素分子梯度的调控。一项研究表明裂殖可能被前部的神经系统抑制,而另一项研究检验了促进其发生的生物力学因素和组织特性。
再生可以通过以实验手段将涡虫切成小段来诱导实现,但是裂殖却缺少稳定可靠的诱导方式,这极大地限制了对这一过程的研究。但是Arnold等人发现,将涡虫转移到食物受限且水不循环的培养环境中,可以在长度超过4毫米或5毫米的涡虫中诱导裂殖(图1)。通过分析影像记录,研究人员发现裂殖持续30分钟左右,并产生出约1毫米长的片段,裂殖发生频率与亲虫大小相关。Arnold等人还发现,当他们在正常培养物中对放置在涡虫上方的盖玻片施加压力时,涡虫会沿其前后轴方向分裂成多个规则间隔的片段。这表明,在成虫中存在预先建立的裂殖平面,其数量随涡虫的大小而变化,而隐藏的分段结构是这种关系的基础。
图1:涡虫中的尺寸依赖型裂殖行为。涡虫可以通过裂殖的方式进行繁殖。在这一过程中,一部分组织从涡虫的身体后部掉落,再生成完整的虫体。Arnold等人检验了裂殖过程中的分子和细胞基础。他们发现裂殖的发生频率与亲虫的尺寸相关。对Wnt信号通路中控制涡虫体长方向组织分布的特定蛋白表达,进行试验干扰(结果未显示),并不影响裂殖的位置,但会增加或降低裂殖事件的频率。作者表明Wnt信号通路可以调节涡虫前端抑制裂殖行为的神经元细胞群的小尺度分布模式(方框内),并表明这些神经元的分布模式随动物大小而变化。
以饥饿法和加压法诱导的涡虫裂殖作为工具,作者研究了哪些分子对于尺寸依赖型裂殖是必不可少的。在一项筛选实验中,他们利用不同的RNA分子来选择性地抑制组织形成过程中的蛋白质,这其中就包括Wnt 和 TGF-β信号通路中的蛋白质[4,6,9,10]。这些针对性的干扰影响了裂殖发生的频率,例如APC是一种抑制Wnt信号通路的蛋白,阻断其表达会导致以伸展行为为标志的连续裂殖尝试频率加倍。然而,干扰这些信号通路并没有影响裂殖平面沿体轴的定位。因此,Wnt 和TGF-β信号通路对裂殖行为的调控似乎与它们在轴向形成中的功能无关。
曾有一项基因表达分析研究揭示了编码Wnt和TGF-β信号通路蛋白的基因与中枢神经系统(CNS)神经细胞的基因存在共表达现象。在Arnold及其同事的研究中,去除涡虫包含头神经节(即共同组成大脑的两个神经元簇)的前端会导致裂殖行为的延迟。作者在一种神经元转录因子蛋白表达被抑制的涡虫中也发现了这种延迟现象,这种转录因子已被证明在CNS形成过程中是必不可少的。
Arnold等人发现了一组对机械刺激敏感的神经元细胞在Wnt和TGF-β信号通路的下游起作用来抑制裂殖行为。作者证明了Wnt和TGF-β信号通路共同调控了这些和其他特定神经元的分布。检验这些轴向分布的关键控制因子是如何控制涡虫神经系统的分布形成,令人感到兴奋——不同种类细胞分布的一个重要问题是祖细胞如何整合这些信号通路来诱导特定种类神经元细胞的再生。
然而Arnold等人将他们的分析集中在裂殖的诱导上,关于释放的组织碎片如何形成完整动物仍知之甚少。例如,目前尚不清楚这些涡虫在裂殖后是否以被切断后的方式再生。在这两种情况下,被称为新胚芽的干细胞群聚集在组织碎片的伤口部位,形成被称为芽基的团块,进而再生不同的器官和组织。但是切割或裂殖平面的位置信息如何传递给新胚芽,目前仍不清楚。
通过裂殖进行无性繁殖的不止是涡虫,也是其它蠕虫样生物(包括阿科尔扁形虫和其他的无腔扁虫和环节动物)进行种群扩张的主要策略,这种裂殖发生在动物的身体后部。海葵也可以通过裂殖来进行无性繁殖,而出芽生殖——一种与裂殖有关的无性繁殖策略——在淡水动物水螅中得到了很好的描述,这种生殖方式与再生密切相关。
在不同的模式生物中对裂殖和出芽生殖进行详尽的研究极为重要,因为在这些过程中,组织器官的形成是在无伤口的情况下完成的,因此可能与受伤后的再生过程不同。如果涡虫裂殖和被切割后的再生过程确实相同,那么未来的研究应该确定缺乏损伤信号的裂殖组织中的补偿机制。这种研究对于理解损伤信号和组织器官形成信号如何配合以启动再生过程至关重要。



